Phagocytes and Phagocytosis
Ultimately, the vast majority of microorganisms are killed by either macrophages or polymorphonuclear leukocytes (PMNs) and the processes involved are part of both the innate and the adaptive immune systems. Phagocytosis is the process by which phagocytes (eater-cells) take-up invading organisms and toxins and subsequently kill them. This happens both spontaneously and with the aid of an opsonin. An opsonin is a chemical that binds to the invading organism; phagocytes have specific receptors on their surface for these opsonins. The most obvious and probably most important example of this is the antibody.
Polymorphonuclear leukocytes (PMNs) (also known as granulocytes) are three cell types that all derive from the lymphoid progenitor cell line. They are:
Neutrophils (so named due to their poor response to staining)
Eosinophils (stains positively with eosin (an orange/red stain))
Basophils (stains positively with basic stains)
Their basic properties are outlined below:
Neutrophils
~90% of the polymorph population
- short lived cell type ~2-3 days
- made in bone marrow
o half life in blood ~ 6 hours
- non-dividing cell in the blood
- function by taking-up and killing microbes
- 3-4 x 109 stored in bone marrow and released into the blood in the event of infection
o 5 fold increase in production during infection
Eosinophils
~10% of PMNs
- short lived cell type ~2-3 days
- known to have bacteriocidal activity
- thought to be important in killing parasitic worms by exocytosis
Basophils
< 1% of PMNs
- life span not known
- functionally very different from other PMNs
o no killing function
o initiator of acute phase
§ releases granules that:
· dilate blood vessels
· increase vessel permeability, and
· are chemoattractants
Macrophages
- Macrophages are produced in bone marrow as pro-monocytes.
- These are released into the blood as monocytes
o Monocytes are thought to be active against microbes to some extent
- After ~ 1 day monocytes migrate into the tissues
- In the tissues they differentiate into macrophages
o – Each tissue macrophage is functionally different, i.e. an alveolar macrophage (lung) is different from a kuppfer cell found in the liver.
o Osteoclasts in the bone are sometimes considered to be macrophages as they derive from monocytes in the same way.
- Macrophages are functionally much more complex then neutrophils:
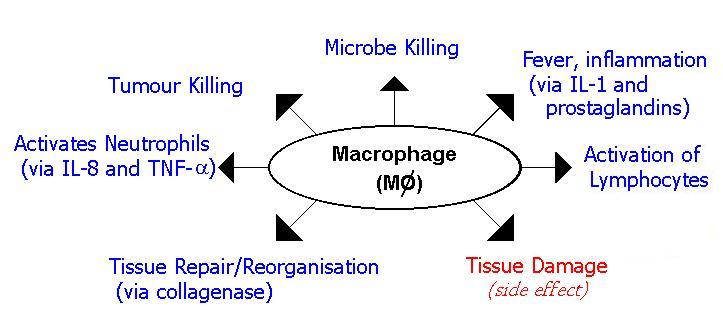
As their name suggests (macrophage = big eater), the first function of macrophages to be identified was killing cells. Whilst this is important, a major aspect of their function is as secretory cells. Macrophages release >100 different types of molecule, some continuously, some during an immune response and others during opsonization.
Lysozyme breaks down the peptidyoglycan wall of bacteria, thereby lysing the cell (hence its name) and is released constantly by macrophages. Lysozyme is found in respiratory secretions and tears. Elastase and collangenase break down elastin and collagen structures in human connective tissue and are responsible for tissue damage in the immune response. These are released during opsonization. The Complement factors C1q, C2, C3, C4, FP and FD are all produced by the macrophage. They are all released during the immune response. (The role of complement is explained in this section). Various inflammatory cytokines are released by the macrophage during the immune response. Included in this list are IL-1, TNF-a, IL-8, IL-6, G-CSF, M-CSF and GM-CSF. The last of these (Granulocyte-Colony Stimulating Factor, Macrophage-Colony Stimulating Factor and Granulocyte/Macrophage-Colony Stimulating Factor) act to stimulate the production of neutrophils, macrophage and both respectively.
Macrophages and Neutrophils
Both macrophages and neutrophils act in essentially the same way but neutrophils are mainly responsible for the killing of acute infective organisms and macrophages for chronic, longer term infections.
The first phagocytic cell to contact an invading microbe is the resident macrophage. It has very poor killing ability (this is due to the way the immune system is tightly controlled) and its main function at this point is to activate the immune response.
Once an immune response is initiated the first cells to infiltrate the site are neutrophils. These may kill the infection. If they do not, then the macrophage becomes more important as the infection becomes chronic. Macrophages infiltrate the site and are activated by T-helper cells. These activated macrophages are phenotypically different.
Neutrophils die almost immediately after killing the ingested bacteria. The debris of dead and dying neutrophils is the major constituent of pus and myeloperoxidase released by the dead cells gives pus its colour. This is useful clinically as green sputum is very indicative (though, not strictly diagnostic) of a bacterial infection. In the advent of a viral infection the body's response is to produce more T-lymphocytes. A differential blood count shows raised white cells whenever there is an infection, if the majority of these are neutrophils it suggests a bacterial cause and if the majority are lymphocytes then the infection is likely to be viral.
Phagocytosis
Phagocytosis is a two step process:
1. Attachment
2. Uptake
Attachment is either via an opsonin or to a common bacterial antigen such as LPS. LPS (Lipo-polysaccharide) is found on the surface of Gram –ve bacteria. It is not found on any host cells and as such is a logical target for phagocytes. The molecule CD14 is found on macrophages and binds specifically to LPS. Other receptors include the mannose receptor, the glucan receptor and CD11b/CD18. All of these receptors bind to bacterial carbohydrates and thus enable to the phagocyte to engulf the bacteria.
Both macrophages and neutrophils have Fc receptors on their surface. Fc means fragment crystallisable and corresponds to the conserved region of an antibody and hence regardless of the antigen-specificity of the antibody the phagocyte can bind to it. Once bound to a pathogen via the specific antigen, the antibody binds to the Fc receptor and this enables the phagocyte to engulf the pathogen. There are several different Fc receptors; they are referred to by Greek letter, i.e. the FcaR binds to the conserved region of the IgA molecule and the FcgR to the IgG antibody etc. This is opsonization. Other molecules that are able to act as opsonins include C3b and C3bi which are by-products of the Complement Cascade. CR3 (Complement Receptor) is found on phagocytes and this binds to C3b C3bi, enabling the pathogen to which they are bound to be opsonized.
Microbes are taken into membrane-bound vesicles that are named phagosomes. Immediately after this the phagosome fuses with another membrane bound vesicle named a lysosome. The fused vesicle is termed a phago-lysosome. The lysosome contains various toxic species that are used to kill the pathogen. They are contained within a separate membrane-bound component so they are not lethal to the phagocyte. At this point the pH falls rapidly to around 4.0, causing the receptor to dissociate from the microbe and be recycled to the cell surface. The low pH also slows microbial growth and favours the enzymes that function to kill the microbe. (Move mouse over image).
Lysosomes
Neutrophils have two distinct populations of lysosomes . One of these contains myeloperoxidase and the other lactoferrin. Macrophages have only one type of lysosome and have neither lactoferrin nor myeloperoxidase.
Respiratory Burst
This process is the primary means of killing bacteria. It involves a series of coordinated changes in the metabolism of oxygen. The point of this is to produce reactive species of oxygen.
The superoxide anion o O2-
Hydroxyl radical o OH
Hydrogen peroxide H2O2
Nitric oxide NO o
All of these are very reactive and will react with the molecules found both on and in any biological cell and in doing so will cause irreparable damage to the cell. This mechanism is very energy-dependent and consumes large amounts of O2 which is why the neutrophils die shortly after engulfing bacteria.
Antigen Presenting Cells
Another important function of phagocytes is as antigen presenting cells. This function is discussed in the section on T-helper cells.